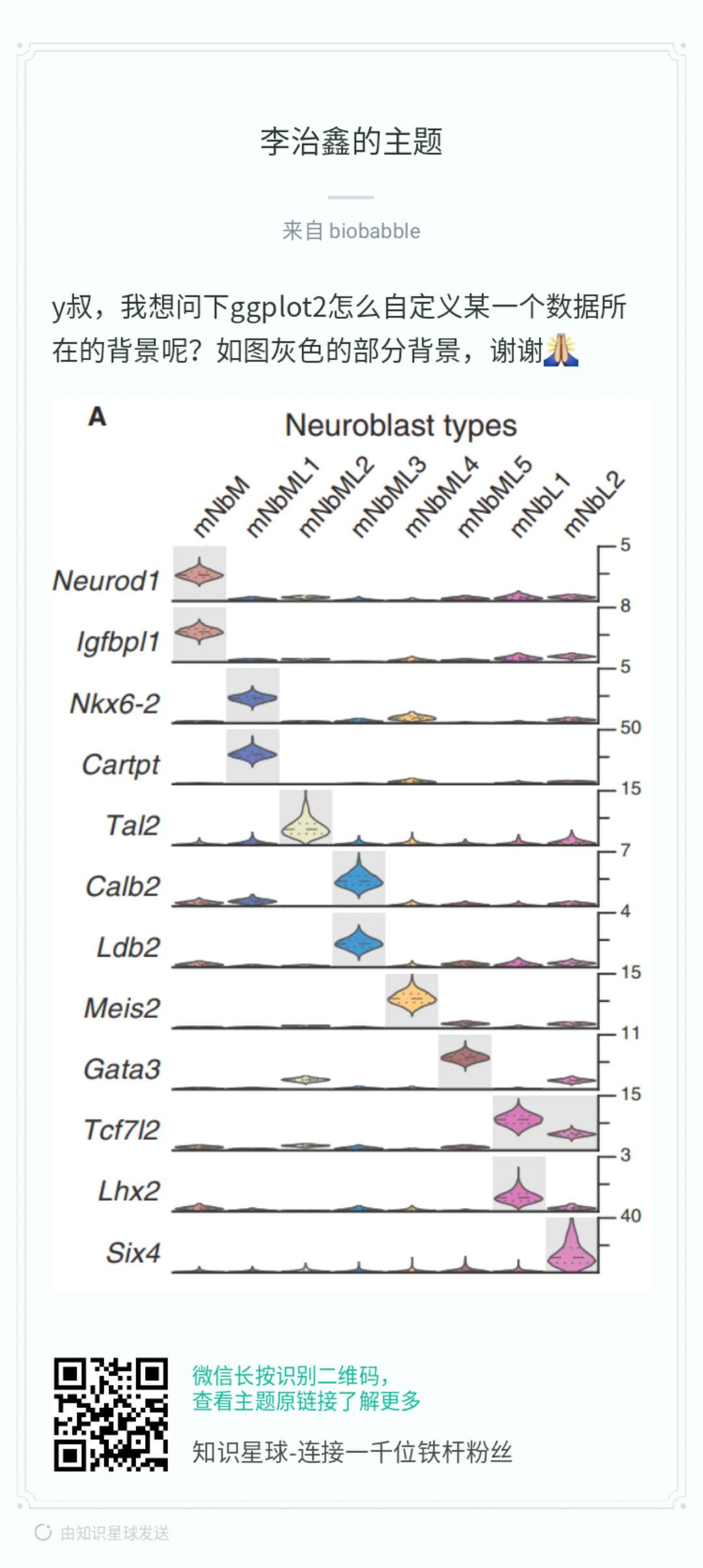
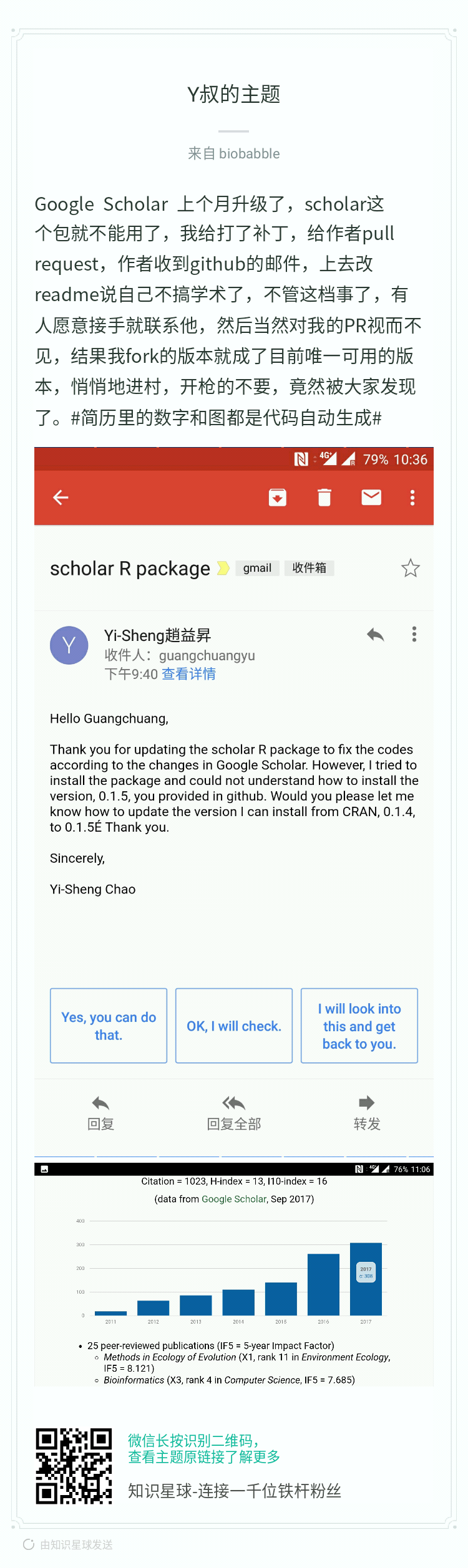
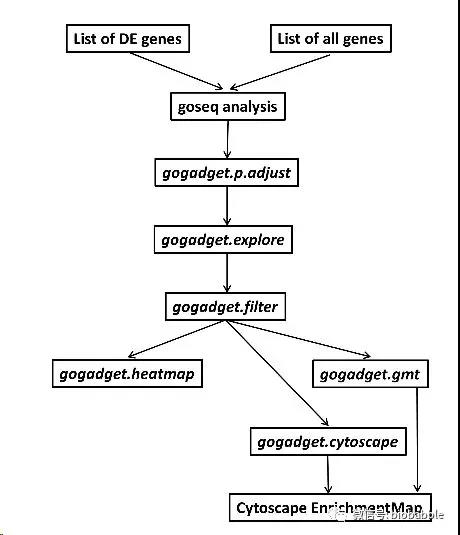
之前有写过读文献的文章,也介绍了如何获取文献,那是我还在读硕士的时候写的,覆盖了找文献、文献管理、下载文献、订阅期刊等方方面面,说到下文献,最大的神器当然是sci-hub,但是sci-hub的域名sci-hub.cc, sci-hub.bz, sci-hub.io纷纷都沦陷,最近国内有一神器geenmedical,整合了影响因子、引用数等指标,然而这一神器的下文献功能只是提供外链,有一种说法是geenmedical=pubmed+scihub,当然geenmedical的资源还可能来自于百度学术、researchGate等,不都是来自于sci-hub,然而我们必须承认的是最大的文献提供来源必须还得是sci-hub,所以如果sci-hub挂了,基于sci-hub的神器也就神奇不起来了。如果有人跟你说忘记sci-hub吧,用geenmedical,那绝对是标题党!这里并没有说geenmedical不好,它整合了多方面的资源并且本地化得非常好,很适合国内使用。
比如我在geenmedical里搜clusterProfiler,给出的下载链接第一个就是sci-hub的:http://doi.org.sci-hub.cc/10.1089/omi.2011.0118,然而这个链接是不可用的!当然它给出了比如百度学术的链接是可以下载到文献的。然而还是那句话,论「海盗精神」我只服sci-hub,虽然我之前也介绍了《sci-hub如果挂了,你还有神器下文献吗?》,但任何下文献的神器,在sci-hub面前都是渣渣。